|
Carnivorous Plant Newsletter
Volume 30, Number 4, December 2001, pages 120 - 125; Back Cover
Technical Refereed Contribution
Heliamphora folliculata,
a New Species of Heliamphora (Sarraceniaceae)
From the ‘Los Testigos’ Table Mountains in the South of Venezuela
Dr. Andreas Wistuba • Mannheim • Germany
Dr. Peter Harbarth • Dossenheim • Germany
Thomas Carow • Münnerstadt • Germany
Keywords: new taxa: Heliamphora folliculata, Venezuela.
Received: 14 April 2001
Introduction
In January 2001 we had the great opportunity to study the flora of some of the least explored of all table mountains of southern Venezuela. Some of the expedition’s destinations were the four table mountains in the Aparaman-group (‘Los Testigos’), consisting of Aparaman Tepui, Murosipan Tepui, Tereke-Yuren Tepui and Kamarkaiwaran Tepui (Steyermark, 1986; George, 1988). The Aparaman-group is located in the northern part of the Guayana Shield, northwest of Ptari Tepui and east of Auyan Tepui. The altitude of these tepuis lies between 2100 meters (Tereke-Yuren) and 2700 meters (Kamarkaiwaran).
On all tepuis of the Aparaman-group we found a very interesting, and previously undescribed species of Heliamphora notable for the unique anatomy of its pitcher-appendage. The appendage is fused with the back of the pitcher forming a hollow structure running downwards on the outside of the back of the pitcher, apparently working as a chamber to store nectar produced by the glands of the appendage. Such a structure has never before been observed in Heliamphora or any other member of Sarraceniaceae.
Heliamphora folliculata Wistuba, Harbarth & Carow spec. nov.
Caudex ramosus; foliis rosulatis; amphoris 20-30 cm longis, tubulosis, basin versus infundibuliformibus, extus glabris, parte superiore interiore glabrio; appendice folliculato amphora deorsum adnato.
Inflorescentiis 3-4-floris, racemosis, ad 35 cm longis; flores nutantes; pedicellis 3-6 cm longis; tepalis 4 oblongo-lanceolatis, albidis vel pallide-roseis, 4-4.5 cm longis; staminibus 10, 1-serialibus, filamentis 8 mm longis; antheris oblongo-lanceolatis, ca. 8 mm longis; ovario valde tomentoso; stylo glabro; stigmate 3 lobato; seminibus fuscis, oblongis, ca. 2-3 mm longis, testa conspicue membranaceo-alata.
Herbaceous perennial; Rhizomes branching, plants forming dense clumps. Pitchers infundibulate in the lower third, cylindrical in the upper two thirds, 20-30 cm long and 5-6 cm wide in the upper part; upper part of the pitchers almost completely glabrous on the inner side; pitchers frequently compressed between the front and the back, the front being pressed inwards, leading to a kidney shaped mouth in most older pitchers when viewed from above; the left and right parts of the lobes forming the pitcher-tube often fused asymmetrically at the front of the pitcher-mouth; older pitchers deep red, younger ones golden-green with red veins; pitchers of younger plants much less compressed and more symmetric. Lid bent sharply towards the front, 1 cm long, 0.5 cm wide and 0.5 cm high, slightly helmet shaped; back of the lid of adult pitchers bearing a hollow structure running downwards 5-10 mm; channel from the hollow chamber to the front of the lid. Inflorescence about 35 cm long, 3-4 flowers, peduncle glabrous; pedicels 3-6 cm long; bracts ovate 3-5 cm long; tepals 4, oblong-lanceolate 4-4.5 cm long, 1.8-2.5 cm wide, white to whitish-pink; 10 stamens in 1 series, filaments 8 mm long, anthers oblong-lanceolate, approximately 8 mm long, 1.5 mm wide; ovary 3 celled, pubescent, style glabrous; seed approximately 2-3 mm long, compressed, ovate, irregularly winged.
Specimens examined
Heliamphora folliculata: Murosipan Tepui, 8 February 2001, Wistuba, Harbarth & Carow No. Mur 08.01.01/1, holotype, flowering specimen (VEN)
Heliamphora folliculata: Murosipan Tepui, 8 February 2001, Wistuba, Harbarth & Carow No. Mur 08.01.01/2, isotype, flowering specimen (VEN)
Heliamphora folliculata: Aparaman Tepui, 8 February 2001, Wistuba, Harbarth & Carow No. Ap 07.01.01/1, isotype, flowering specimen (NY)
Distribution
Heliamphora folliculata is only known from the Aparaman-group of Tepuis, where we found it on all four mountains: Aparaman Tepui, Murosipan Tepui, Tereke Tepui and Kamakeiwaran Tepui. The largest population was observed on Aparaman Tepui where the plants were growing in dense overhanging populations, partly on vertical rock-faces. We did not observe notable morphological differences among the four populations, except for a single group of plants on Kamarkeiwaran that showed an extremely short appendage.
The two other tepuis in the area (Auyan Tepui and Ptari Tepui) are well explored. They are known to house two distinct species (Heliamphora minor on Auyan Tepui, and Heliamphora heterodoxa on Ptari Tepui) (Maguire, 1978; Steyermark, 1984). Differences between these two species and Heliamphora folliculata are given in Table 1. Accordingly, we believe that Heliamphora folliculata does not occur on these tepuis, and instead is endemic to the table-tops of the Aparaman-group.
Table 1: Comparison between Heliamphora folliculata, Heliamphora minor and Heliamphora heterodoxa.
|
Heliamphora folliculata |
Heliamphora minor |
Heliamphora heterodoxa |
Pitcher: |
|
|
|
Dimensions |
20-30 cm long
5-6 cm wide |
8-20 cm long
3-8 cm wide |
15-40 cm long
5-6 cm wide |
Shape |
Infundibulate in the lower third, cylindrical in the upper two thirds;
Pitchers frequently compressed between the front and the back, the front being pressed inwards |
Slightly ventricose in the lower part, narrowly expanded to tubular in the upper part |
Infundibulate to ventricose in the lower third, slightly infundibulate in the upper part, expanded near the mouth |
Lid: |
|
|
|
Dimensions |
1 cm long
0.5 cm wide |
0.5-1 cm long
0.3-0.5 cm wide |
1-3.5 cm long
1–3 cm wide |
Shape |
Bent sharply towards the front, slightly helmet- shaped; back of the lid of adult pitchers bearing a hollow structure running downwards 5-10 mm; not constricted at the base |
Strongly helmet-shaped,
extremely constricted to stalked at the base |
Flattened to helmet-shaped;
only slightly constricted around the base |
Anthers/ Stamens : |
|
|
|
Length |
8 mm |
4 mm |
6-8 mm |
Number |
10 |
15 |
8-14 |
Ecology
Heliamphora folliculata plants were usually found growing in exposed areas with short vegetation such as grasses, bromeliads, Xyris (Xyridaceae) or Stegolepis (Rapateaceae). These plants do not shade the Heliamphora significantly, in fact we never found plants growing in shady locations. Most of the dense patches of Heliamphora folliculata were found on steep rocks with good drainage. Only rarely have we observed H. folliculata growing at really wet places, such as near shallow ponds or seepage slopes.
We opened a number of pitchers but a significant amount of prey was not present, so the efficiency of the trapping mechanism remains doubtful for at least during the dry season (the time we visited the Aparaman group). We have made similar observations for other Heliamphora species on other tepuis. However, other authors have reported an efficient capture rate of insects (Gonzales et al., 1991; Jaffe et al., 1992). Jaffe (1992) reported ants (Formicidae) as being a major component of the captured arthropods in Heliamphoranutans pitchers. For different Heliamphora species, they observed approximately 5-40 arthropods captured by a single leaf.
Another observation associated with the trapping mechanism is the fact that most of the nectar chambers of the appendages were opened and thus destroyed, most likely by wasps seeking the nectar inside. Unfortunately we had no opportunity to observe this happening, so we could not confirm wasps were responsible for this damage.
Interestingly, we frequently observed big black bumblebees pollinating the flowers. Bumblebees have been reported to be pollinators for other species of Heliamphora (Renner, 1989).
Another interesting observation was that there was a high percentage of pitchers damaged, most likely by caterpillars.
Related species
The unique architecture of this species’ lid places it in an isolated position within the genus. H. folliculata does not seem to be particularly closely related to any species in the genus. The lid also easily distinguishes H. folliculata from H. heterodoxa and H. minor (the two Heliamphora species that occur on neighboring tepuis).
Etymology
The name ‘folliculata’ was chosen to highlight the ‘bubble’ (=follicle) formed by the nectar spoon, the most distinct characteristic of this species.
Discussion
We assume that the structure formed by the lid is a highly derived characteristic rather than a relict lost in all other species. The diversity of the pitcher nectar spoon in the various species of Heliamphora--H. minor which has a highly differentiated helmet-like structure, H. tatei which has a rather simple flag-like structure, H. heterodoxa var. exappendiculata with its rudimentary lid, and the new species H. folliculata as described in this paper--illustrate the various ways the nectar spoon is shaped in order to function as the predominant structure for the attraction of prey (Jaffe et al., 1995). While other authors reported significant amounts of prey (Gonzales et al.; 1991; Jaffe et al., 1992), we view the efficiency of prey capture to be very poor when compared to the amount of dead arthropods regularly found inside the pitchers of other carnivorous pitcher-plants such as Nepenthes or Sarracenia. However, in the habitats where these latter plants grow, the competition with other non-carnivorous plants is much higher, and also many more arthropods occur there. So it is possible that the true prey-capture efficiency of Heliamphora is much higher than it appears at first sight if the extremely low overall density of the arthropod-populations on the table-tops of the tepuis is taken into consideration. The elaborate structures of the nectar spoons might facilitate the capture of a few more prey, and these small numbers might well be sufficient to give the plant an advantage over other non-carnivorous plants in this harsh environment.
Literature
George, U. 1988, Inseln in der Zeit, GEO im Verlag Gruner + Jahr AG & Co., Hamburg.
Gonzalez, J.M., Jaffe, K., and Michelangeli, F. 1991, Competition for Prey Between the Carnivorous Bromeliaceae Brocchinia reducta and Sarraceniaceae Heliamphora nutans, Biotropica 23 (4 Part B): 602-604.
Jaffe, K., Blum, M.S., Fales, H.M., Mason, R.T., and Cabrera, A. 1995, On Insect Attractants from Pitcher Plants of the Genus Heliamphora (Sarraceniaceae), Journal of Chemical Ecology, 21(3): 379-384.
Jaffe, K., Michelangeli, F., Gonzalez, J.M., Miras, B., and Ruiz, M.C. 1992, Carnivory in Pitcher Plants of the Genus Heliamphora (Sarraceniaceae), New Phytologist 122(4): 733-744.
Maguire, B. 1978, Sarraceniaceae (Heliamphora), in: The Botany of the Guyana Highland Part-X, Memoirs of the New York Botanical Garden 29: 36-61.
Renner, S.S. 1989, Floral Biological Observations on Heliamphora tatei Sarraceniaceae and Other Plants from Cerro de la Neblina in Venezuela. Plant Systematics & Evolution 163 (1-2): 21-30.
Steyermark, J.A. 1984, Venezuelan Guyana, Annals of the Missouri Botanical Garden 71: 302-312.
Steyermark, J.A. 1986, Expedition to the Lost World, Annals of the Missouri Botanical Garden 74(7): 5-7.
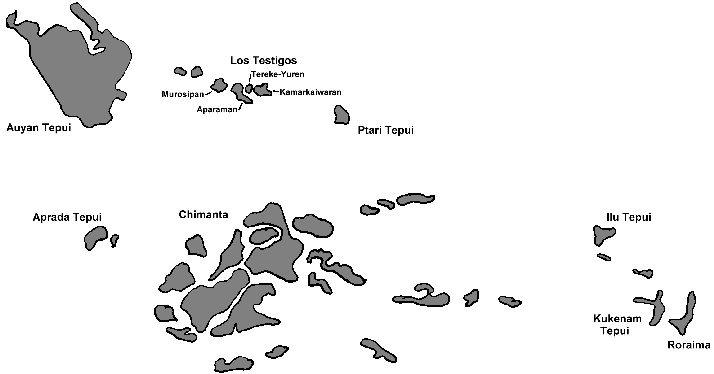
Figure 1: Map indicating the peaks in the complex of tepuis. The indicated peaks have elevations of more than 1500 meters higher than the surrounding plains. Prepared by Dr. Peter Harbarth and Dr. Andreas Wistuba.
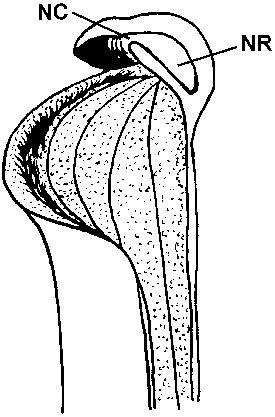
Figure 2: Drawing of the top of a H. folliculata pitcher in cross section. “NC” indicates the nectar channel under the lid appendage, “NR” indicates the nectar reservoir. Drawn by Dr. Peter Harbarth.

Figure 3 A group of H. folliculata pitchers on Aparaman Tepui. Photograph by Dr. Andreas Wistuba.

Figure 4: Details of H. folliculata pitchers. A)nectar (dark red) running down from the lid, B)lid showing the hollow hunchback-like structure, C)pitcher compressed front to back, leading to kidney shaped mouth, D)nectar reservoirs opened by insects. Photographs by Dr. Andreas Wistuba.

Back Cover: A new species of Heliamphora, photographed by Dr. Andreas Wistuba.
|


