|
Carnivorous Plant Newsletter
Volume 32, Number 3, September 2003, pages 79 - 92; Front Cover
Technical Refereed Contribution
Four New Species of Sundews, Drosera
(Droseraceae), from Brazil
Fernando Rivadavia • São Paulo • Brazil
Keywords: new taxa: Drosera tentaculata, Drosera grantsaui, Drosera camporupestris, Drosera viridis, Brazil.
Received: 27 December 2002
Abstract
Four new species are described from Brazil for the sundew genus Drosera (Droseraceae): Drosera tentaculata F.Rivadavia spec. nov., D. grantsaui F.Rivadavia spec. nov., D. camporupestris F.Rivadavia spec. nov., and D. viridis F.Rivadavia spec. nov. The morphological characters distinguishing these from other similar or related taxa are discussed together with habitat information, detailed illustrations, and distribution maps. Drosera grantsaui and D. viridis are included in subgen. Drosera sect. Drosera, whilst D . tentaculata and D. camporupestris are included in subgen. Drosera sect. Oosperma. Furthermore, D. chrysolepis Taubert and D. graminifolia A.Saint-Hilaire are moved from sect. Drosera to sect. Oosperma.
Introduction
The sundews of the genus Drosera comprise nearly 150 species spread worldwide, mostly in the Southern Hemisphere and especially in southwestern Australia (Diels, 1906; Schlauer, 1996). Around fifteen Drosera species are presently known to occur in Brazil (Diels, 1906; Silva & Giulietti, 1997), the first of which were published by A.Saint-Hilaire (1824a; 1824b).
Initiated by this author in 1990, ongoing field studies with Drosera spp. throughout Brazil have resulted in the discovery of four new species, all of which were previously mistaken for other similar or related taxa. However, upon closer examination each of these new species is clearly distinctive from all other Brazilian Drosera taxa in their vegetative, floristic, and ecological characteristics. These new taxa and other native Brazilian Drosera species were cultivated in a greenhouse in São Paulo over a five year period, during which observations and comparisons were carried out.
Results
Drosera tentaculata F.Rivadavia, spec. nov. TYPE -- BRAZIL . Minas Gerais: Município de Diamantina, no alto da Cachoeira dos Cristais, estrada para Biribiri, c .1000 m alt., 31/Jul./2002, Rivadavia & Gibson 1376 ( holotype -- SPF). Figure 1:A-J.
Drosera montana A.Saint-Hilaire var. tomentosa (A.Saint-Hilaire) Diels affinis sed plantis frequenter columnas brevibus e foliis defunctis compactis formantibus, margine laminae apice glandulis 8-12, 2-9 mm longis, rectangularibus, longe stipitatis obsita; foliis obovato-cuneatis; scapibus 3.5-18.5 cm longis (inflorescentia inclusa); inflorescentia 1-9-flora; seminibus ovoideo-ellipsoideis.
Perennial herbs with leaves in rosettes flat on the ground or atop short columns of dead leaves up to 2.5 cm high. Stipules 2-4 mm X 0.8-1.5 mm, translucent white in color, the upper half to two-thirds divided into 7-11 filaments, stipule filaments shorter towards the center of the stipules. Leaves 3-13(-17) mm long excluding retentive glands, obovate-cuneate (cuneate), orange-green to deep wine-red in exposed habitats, green in shadier habitats; petioles 0.5-4(-8) mm X 0.8-1.5 mm, usually greener than the lamina, sparsely eglandular-pilose to glabrous adaxially, abaxially sparsely to densely covered with shaggy white eglandular hairs up to 1 mm in length; lamina 2-9 mm X 1.5-4(-8) mm, more or less eglandular-pilose abaxially, especially near the base and along the margins, adaxially covered with numerous red retentive glands, including 8-12 red, horizontally flat, retentive glands on the apical margin, 2-9 mm long, each with a narrowly-obcuneiform stalk and a linearly-rectangular head. Inflorescences 1-3 per plant, 3.5-18.5 cm long including scape, with 1-9 flowers each, erect, never bifurcating, glandular-pilose especially towards the apex, eglandular-pilose on lower one-third of the scape, especially towards the base; pedicels 2-5 mm long, inserted 2-20 mm apart, glandular and often eglandular-pilose; sepals 5 united for 1/3 to 1/2 of length, 2-4 mm X 0.7-1.5 mm, oblong-ovate, abaxially glandular and often eglandular-pilose; petals 5.5-8.5 mm X 3-4 mm, obovate, dark pink-lilac in color; ovary 1-1.7 mm long at anthesis, trilobed, globose, becoming ellipsoid in fruit, 2-2.5 mm long; carpels 3; styles 3, 2-4 mm long, stigmata indistinct, basally bifurcate, thickening somewhat towards the apex with several constrictions or without constrictions or with distinct stigmata inconspicuously flabellate to bifurcate; stamens 5, 2.5-4 mm long; seeds black, 0.3-0.45 mm long, black, ovoid-ellipsoid, reticulate. Roots fine and fibrous.
Drosera tentaculata is placed in subgen. Drosera sect. Oosperma sensu Schlauer (1996). It is easily distinguishable by its obovate-cuneate leaves with 8-12 large apical retentive glands 2-9 mm long, each with a narrowly-obcuneiform stalk and a linearly-rectangular head. Such tentacles are only found in one other New World species, D . sessilifolia A.Saint-Hilaire, which differs in its annual habit, yellowish to orange-reddish cuneate leaves, 5 carpels, 5 stigmata (without basal bifurcations), and ovoid seeds.
The styles with several constrictions and indistinct stigmata found on D. tentaculata from the Serra do Cipó are unique in the genus Drosera. Yet this character is apparently variable in D. tentaculata since the styles of plants from the Serra do Cabral do not have constrictions while those from the Diamantina region have distinct stigmata varying from inconspicuously flabellate to bifurcate. Such variability in style and especially stigmata morphology has also been observed in other Drosera spp. in Brazil, especially D. montana var. montana.
Also variable in D. tentaculata is the amount and distribution of eglandular hairs on the inflorescences, which are more frequent near the base and apex of the inflorescence but are often almost completely absent. The number and distribution of eglandular hairs have even been observed to vary on different inflorescences belonging to the same plant.
Drosera tentaculata is most commonly confused with D. montana var. tomentosa due to similiarities in overall shape and size, inflorescence pubescence, and to the fact that both taxa often occur sympatrically. Other than the absence of the large apical retentive glands, D. montana var. tomentosa can easily be distinguished by its oblongo-obovate leaves; on average longer petioles; inflorescences up to 33 cm in length, often bifurcate, with as many as 22 flowers each; and seeds cuneate-ellipsoid.
This new species is commonly found growing near or sympatrically with other Drosera spp. such as D. camporupestris F.Rivadavia, D. chrysolepis, D. graminifolia, D. montana var. montana, D. m. var. schwackei Diels, and D. m. var. tomentosa. No hybrids between any of these and D. tentaculata have ever been observed, including the morphologically more similar taxa of the D. montana-complex. Drosera tentaculata occurs in “campo rupestre” vegetation, usually in areas that suffer intense desiccation during the dry season, which is when flowering usually occurs (July to October). It is found predominantly among sparse grasses in fine white sand mixed with white quartz gravel, or in sand mixed with black humus, or even on bare sandstone surfaces. The rosettes often lose vigor after flowering, growing slowly over the following few months.
Although it is apparently better suited to survive in relatively dry habitats, D. tentaculata has been found growing in very boggy soil at one location, where the leaves were exceptionally large and cuneate in shape (Rivadavia & Gibson 1402 and Rivadavia 1591--SPF). These unusual measurements are included between parenthesis in the above description of this new taxon.
Drosera tentaculata is endemic to the Cadeia do Espinhaço Highlands in the states of Minas Gerais and Bahia, where it has been collected on the Pico das Almas (c.1500 m elevation), Serra do Cabral (c.1280 m elevation), and Serra do Cipó (c.1250-1400 m elevation) as well as on highlands near the towns of Diamantina (c.950-1400 m elevation), Gouvea (c.1260 m elevation), Milho Verde (c.1100 m elevation), and Itacambira (c.1200-1350 m elevation). (Figure 5)
The epithet “tentaculata” refers to the exceptionally large tentacle-like retentive glands present on the apex of the leaves of this new species.
Paratypes -- BRAZIL. Bahia: Rio de Contas, lower NE slopes of the Pico das Almas, 17/Feb./1977, Harley et al. 19568 (SPF). Minas Gerais: Diamantina, estrada p/ Medanha, 12/Oct./1984, Mello-Silva et al. 5379 (SPF); Diamantina, cachoeira dos Cristais, 7/Jul./1995, Rivadavia & Pinheiro 451 (SPF); Diamantina, estrada p/ Curvelo, 27/Feb./1997, Rivadavia & Pinheiro 571 (SPF); Diamantina, estrada p/ Tapetes Arraiolos, 27/Feb./1997, Rivadavia & Pinheiro 580 (SPF); Diamantina, estrada p/ Tapetes Arraiolos, 10/Jul./1999, Rivadavia, et al. 1106 (SPF); Diamantina, cachoeira dos Cristais, 31/Jul./2002, Rivadavia & Gibson 1376 (SPF); Gouvea, estrada Diamantina-Curvelo, 2/Aug./2002, Rivadavia & Gibson 1402 (SPF); Gouvea, estrada Diamantina-Curvelo, 6/Apr./2003, Rivadavia 1591 (SPF); Itacambira, estrada p/ M.Claros, 5/Mar./1997, Rivadavia 614 (SPF); Itacambira, estrada p/ M.Claros, 14/Jul./1999, Rivadavia & Pinheiro 1143 (SPF); Itacambira, estrada p/ M.Claros , 29/Jul./2002, Rivadavia & Gibson 1364 (SPF); Jaboticatubas, km 142 rodovia L.Santa-C.M.Dentro, 27/May/1972, Joly et al. 2111 (SP); Jaboticatubas, estrada da usina, 21/Aug./1972, Joly & Semir 3067 (SP); Jaboticatubas, km 138-9 rodovia L.Santa-C.M.Dentro, 8/Sep./1972, Joly & Müller 3433 (SP); Jaboticatubas, 10/Apr./1980, Barros 200 (SP); Jaboticatubas, Serra do Cipó, trilha p/ mirante do canyon Bocaina, 5/Jun./1994, Rivadavia 289 (SPF); Jaboticatubas, Serra do Cipó, Fazenda da Serra do Cipó, 11/Sep./1994, Rivadavia 305 (SPF); Jaboticatubas, Serra do Cipó, estrada p/ o mirante do canyon Bocaina, 23/Feb./1996, Rivadavia & Mullins 541 (SPF); Jaboticatubas, Serra do Cipó, km 112 ou 113 da estrada p/ C.M.Dentro, 9/Jul./1999, Rivadavia, et al. 1096 (SPF); Joaquim Felício, alto da Serra do Cabral, 7/Mar./1997, Rivadavia 629 (SPF); Santana do Riacho, caminho da Lapinha, 11/Feb./1991, Arbo et al. 4851 (SPF); Santana do Riacho, rodovia B.Horizonte-C.M.Dentro, 6/Sep./1980, Forero et al. 7762 (SPF) (mixed collection with D. montana var.tomentosa); Santana do Riacho, 2.5 km após acampamento Serra Morena, 28/Jul./2002, Rivadavia & Gibson 1362 (SPF); Serro, Milho Verde, trilha p/ cachoeira do Arco-Íris, 5/Apr./2003, Rivadavia & Neves 1561 (SPF); Serro, Milho Verde, planície ao lado da cidade, 5/Apr./2003, Rivadavia & Neves 1570 (SPF).
Drosera grantsaui F.Rivadavia, spec. nov. TYPE -- BRAZIL. Minas Gerais: Município de Grão Mogol, morro do Jambeiro, nascente brejosa na base do morro, c.7 km da cidade pela estrada para Montes Claros, c.700 m alt., 24/Sep./1994, Rivadavia 299 (holotype -- SPF). Figure 2:A-L.
Drosera communis A.Saint-Hilaire similis sed caulibus 1-28 cm longis; stipulis aeneus; foliis spatulato-linearibus; petiolo 2-12 X 0.2-0.5 mm, villoso; lamina 2-11 X 0.5-1.5 mm, oblongo-lineari, facie inferiore pilis simplicibus sparsioribus obtectis; inflorescentia erecta; seminibus cuneato-fusiformibus.
Perennial herbs with delicate stems 1-28 cm long beset with the deflexed remains of older leaves. Stipules 2-6 X 0.3-1.5 mm at the base, translucent bronze-colored, the apical 2/3 to 1/3 divided into 7-15 filaments shorter towards the stipule center, or stipule with longer lateral filaments and weakly divided into two filament-bearing lobes in the center . Leaves 4-23 mm long, spatulate-linear, alternate on stems, reddish-green or pinkish-red in exposed habitats, green in shadier habitats; petioles 2-12 X 0.2-0.5 mm, sparsely eglandular-pilose; lamina 2-11 X 0.5-1.5 mm, oblong-linear, sparsely eglandular-pilose abaxially, covered with numerous red retentive glands adaxially. Inflorescences 1-3, erect, not bifurcating, sparsely covered with minute glands throughout, especially towards the apex, 3-26.5 cm long including scape, with 1-5 flowers each; pedicels 2-9 mm long, glandular-pilose, inserted 4-15 mm apart; sepals 5 united at basal 1/4 of length, 1.5-4.5 X 0.5-1 mm, oblong-ovate, abaxially glandular-pilose with occasional simple hairs; petals 5, 2-4 X 2-3 mm, obovate, varying from light pink to white to light lilac; ovary 1-1.5 mm long at anthesis, hexalobed or trilobed or smooth, ellipsoid or globose or ovoid, when in fruit ellipsoid, 2 mm long; carpels 3; styles 3, 1.7-2.2 mm long, each basally bifurcate, stigmata flabellate; stamens 5, 1.6-2.8 mm long; seeds dark brown, 1-1.5 mm long, cuneate-fusiform, inconspicuously reticulate, translucent. Roots fine and fibrous.
Drosera grantsaui is included in subgen. Drosera sect. Droserasensu Schlauer (1996). It is distinguished by its conspicuous stems 1-28 cm long, erect inflorescences, spatulate-linear leaves, oblong-linear lamina (eglandular-pilose abaxially), and cuneate-fusiform seeds. Drosera grantsaui is reminiscent of a diminutive D. chrysolepis, but the latter is larger overall, more densely covered with long shaggy white hairs, and has narrowly ellipsoid-ovoid seeds 0.7 mm in length. The recently described D. peruensis T.Silva & M.D.Correa (Silva & Correa, 2002) has similarly shaped leaves but differs from D. grantsaui in its short stocky stems 2-4 cm long, inflorescences 4-7 cm long with eglandular hairs and bearing 2-4 flowers, sepals with eglandular hairs, and seeds oblong c. 4mm long.
Drosera grantsaui is most often mistaken for D. communis (e.g. Silva & Giulietti, 1997) not only because these two taxa often occur sympatrically but also due to the similarity in leaf length, their delicate glandular-pilose inflorescences with few flowers each, and seed shape. Drosera communis (Figure 4: A-G) is distinguished by its stemless rosettes (except when growing underwater) with patent to semi-erect leaves; petioles up to 1 mm wide; obovate to rotundate lamina up to 4 mm wide, glabrous abaxially; stipules translucent white in color; scapes conspicuously ascending at the base; and seeds fusiform. No intermediate specimens have ever been observed in habitats where D. grantsaui and D. communis are sympatric, suggesting that these two taxa are not interfertile. However, rigorous experimental cross pollination experiments have not yet been attempted.
Although most similar to D. communis, D. grantsaui may in fact be more closely related to D. montana var. tomentosa with which it hybridizes at most locations where these two species occur in the same or in neighboring habitats in the states of Bahia, Goiás, and Minas Gerais. Although the hybrid is apparently infertile, unexpectedly it is often found in large numbers. It most closely resembles D. m. var. tomentosa, differing however in its semi-erect leaves only up to 3 mm wide and petioles 3-10 mm long.
Drosera grantsaui is surprisingly widespread and common throughout eastern and central-western Brazil, ranging nearly 2000 km east to west and 1500 km north to south. It is very uniform across this range with only slight variations in overall size, especially stem and leaf length, and in the density of eglandular pubescence on the leaves--all of which can be attributed to ecological or weather conditions, including even the petal coloration which has been observed to vary from year to year in a same population for D. grantsaui, D. communis and D. villosa A.Saint-Hilaire.
Drosera grantsaui apparently has no definite flowering period and can be found in bloom year-round, although it is most common to find plants in bloom late in the dry season, between August and November. This species is found in sandy or peaty soils among sparse to dense grasses in perennially humid to wet habitats, such as seepages or along streams and small rivers. It occurs between 550-1550 m elevation on the Brazilian central highlands, in campo rupestre vegetation on sandstone formations or in areas of “cerrado” vegetation (Brazilian savanna) along “veredas” (stands of Mauritia flexuosa L., Palmae, which grow in grass-covered natural springs). In Brazil D. grantsaui is known from the states of Bahia, Goiás, Distrito Federal, Mato Grosso, Mato Grosso do Sul, Minas Gerais, Paraná, São Paulo, and Tocantins. A single collection is known from Caaguazú Department in south-central Paraguay ( Casas & Molero 6431-- NYBG). (Figure 5)
The epithet “grantsaui ” is in honor of the biologist Rolf Grantsau (see acknowledgements) .
Paratypes -- BRAZIL. Bahia: Abaíra, Catolés, Campo da Mutuca, 23/Mar./1992, Stannard & Silva 52792 (SPF) (mixed collection with D. communis); Abaíra, Catolés, Campo da Mutuca, 23/Mar./1992, Stannard & Silva 52793 (SPF) (mixed collection with hybrid D. grantsaui X D. montana var. tomentosa); Abaíra, Catolés, trilha para o Campo da Mutuca, 29/Jul./1995, Rivadavia & Ganev 491 (SPF); estrada Bahia-S.Domingos, córrego Contagem, 21/May/1983 (UFG); Piatã, Serra do Santana, 27/July/1995, Rivadavia & Ganev 485 (SPF). Distrito Federal: Parque Municipal do Gama, 3/Sep./1964, Irwin & Soderstrom 5900 (UB); Parque Municipal do Gama, 12/Nov./1965, Irwin et al. 10222 (UB); Parque Municipal do Gama, 21/Mar./1966, Irwin et al. 14143 (UB); Parque Municipal do Gama, 10/Apr./1971, Sastre 1134 (UB). Goiás: Alto Paraíso de Goiás, córrego Santana, 1/Feb./1993, Rivadavia 190 (SPF); Alto Paraíso de Goiás, córrego Cristal, 2/Feb./1993, Rivadavia 195 (SPF); Alto Paraíso de Goiás, estrada Brasília-A.Paraíso, 15/Apr./1995, Rivadavia & Ogassavara 390 (SPF); Alto Paraíso de Goiás, Chapada dos Veadeiros, estrada A.Paraíso-Teresina de Goiás, 19/Mar./1997, Rivadavia 655 (SPF); Between Rio Torto and Paranau, 26/May/1895, Glaziou 21121 (P); Caiapônia, cachoeira do Sereno, 30/Oct./2001, Rivadavia 1306 (SPF); Mineiros, Parque Nacional das Emas, rio Formoso, 27/Jul./1991, Rivadavia & Marino 19 (SPF); Mineiros, Parque Nacional das Emas, córrego Avoador, 28/Jul./1991, Rivadavia 21 (SPF); Mineiros, Parque Nacional das Emas, rio Formoso, 4/May/1999, Rivadavia & Sato 1060 (SPF); São Domingos, Parque Estadual de Terra Ronca, 3/Nov./2002, Rivadavia, et al. 1433 (SPF). Mato Grosso: Chapada dos Guimarães, Colégio Buriti, 15/Oct./1973, Prance et al. 18979 (mixed collection with D. communis) (NYBG); Chapada dos Guimarães, próximo ao Colégio Buriti, 29/Apr./1995, Rivadavia & Cardoso 422 (SPF); Vila Bela da Santíssima Trindade, Chapada dos Parecis, 27/Jul./1986, Emmerich et al. 6044 (R). Mato Grosso do Sul: Costa Rica, salto da Rapadura, 28/Oct./2001, Rivadavia 1300 (SPF). Minas Gerais: Botumirim, Serra da Canastra, 19/Nov./1992, Mello-Silva et al. 677 (SPF); Botumirim, Serra da Canastra, 21/Dec./1994, Rivadavia 339 (SPF); Botumirim, Serra da Canastra, 13/Oct./2001, Rivadavia 1271 (SPF); Diamantina, garimpo da antiga Lagoa Azul, 28/Feb./1997, Rivadavia & Pinheiro 583 (SPF); Diamantina, estrada p/ Tapetes Arraiolos, 13/Jul./1999, Rivadavia & Pinheiro 1129 (SPF); Diamantina, estrada que sai de Tapetes Arraiolos, 31/Jul./2002, Rivadavia & Gibson 1370 (SPF); Diamantina, estrada p/ Biribiri, 31/Jul./2002, Rivadavia & Gibson 1378 (SPF); Gouvea, estrada Diamantina-Curvelo, 2/Aug./2002, Rivadavia & Gibson 1400 (SPF); Gouvea, estrada Diamantina-Curvelo, 6/Apr./2003, Rivadavia 1590 (SPF); Grão Mogol, trilha da Tropa, 2/Jun./1994, Rivadavia 279 (SPF); Grão Mogol, morro do Jambeiro, 3/Jun./1994, Rivadavia 285 (SPF); Grão Mogol, trilha da Tropa, 7/Sep./1994, Rivadavia 293 (SPF); Grão Mogol, morro do Jambeiro, 12/Oct./2001, Rivadavia 1261 (SPF); Itacambira, SW da cidade, 18/Dec./1994, Rivadavia 328 (SPF); Grão Mogol, estrada p/ M.Claros, 16/Dec./1994, Rivadavia311 (SPF); Grão Mogol, estrada p/ M.Claros, 5/Mar./1997, Rivadavia 601 (SPF); Grão Mogol, estrada p/ M.Claros, 14/Jul./1999, Rivadavia & Pinheiro 1134 (SPF); Grão Mogol, estrada p/ M.Claros, 13/Oct./2001, Rivadavia 1290 (SPF); Jaboticatubas, Serra do Cipó, Fazenda da S.Cipó, 24/Feb./1996, Rivadavia & Mullins 553 (SPF); Joaquim Felício, alto da Serra do Cabral, 6/Mar./1997, Rivadavia 618 (SPF); Moeda, Serra da Moeda, 12/Mar./2002, Rivadavia 1342 (SPF); Serro, estrada Milho verde p/ Diamantina, 6/Apr./2003, Rivadavia 1585 (SPF); São Roque de Minas, Parque Nacional da Serra da Canastra, 2/Apr./1999, Rivadavia & Peixoto 884 (SPF); Serranópolis, trilha que atravessa a Cadeia do Espinhaço, 17/Jul./1999, Rivadavia & Pinheiro 1151 (SPF). Paraná: Jaguariaíva, rio Cajurú, 24/Apr./1995, Rivadavia 410 (SPF); Sengés, rio Funil, 24/Apr./1995, Rivadavia 413 (SPF). São Paulo: Itararé, rio Verde, 24/Apr./1995, Rivadavia 418 (SPF). Tocantins: Dianópolis, rio Manoel Alvinho, July/1951, Costa 171 (ALCB). s/loc, s/d, Armond 62532 (R). PARAGUAY. Caaguazú: Yhú, cerca y al sur de Yhú, 22/Feb./1982, Casas & Molero 6431 (NYBG) (mixed collection with D. sp. aff. cayennensis Sagot ex Diels).
Drosera camporupestris F.Rivadavia, spec. nov. TYPE -- BRAZIL. Minas Gerais: Jaboticatubas, Serra do Cipó, em campos rupestres dentro da Fazenda da Serra do Cipó, 19°17.597’S 43°34.829’W, 1215 m de altitude, 6/Jul./1995, Rivadavia 447 (holotype -- SPF). Figure 3:1A-F.
Drosera chrysolepisaffinis sed caulibus 5-30 X 3-9 mm; stipulis 7-13 X 2-6 mm, cuneatis apice in filamenta brevia pauca divisis; modo 1-2(3) foliis vivis; petiolis 30-105 X 0.5-1 mm; laminis 10-50 mm longis; seminibus anguste ovoideo-ellipsoideis subtiliter reticulato-foveolatis, 0.6 mm longis.
Perennial herbs with short stems 5-30 X 3-9 mm. Stipules 7-13 X 2-6 mm, translucent bronze-colored, triangular, sometimes apically fimbriate. Leaves 3.5-14 cm long, erect, 1-2(3) functional per plant, several decumbent and dead, deep wine-red in color with green petiole bases in exposed habitats, green in shadier habitats; petioles 25-105 X 0.3-1.5 mm, abaxially and adaxially eglandular-pilose especially towards the base; lamina 10-50 X 1-3 mm, lanceolate, abaxially eglandular-pilose, adaxially covered with numerous red retentive glands up to 6 mm long, the longest located at the base and apex of the lamina. Inflorescences 1-2, erect, occasionally bifurcate at the apex, densely eglandular-pilose, 8.5-23 cm long including scape, with 1-14 flowers each; pedicels 1-4 mm long, inserted 4-16 mm apart, densely eglandular-pilose; sepals 5 united at basal 1/3 of length, 4-7 X 0.5-1 mm, oblong-ovate, densely eglandular-pilose and sparsely glandular-pilose abaxially; petals 5, 6-9 X 3-5 mm, obovate, light to dark pink-lilac colored, occasionally darker at the base; ovary 2 mm long at anthesis, smooth to trilobed, ellipsoid, growing to 3-4 X 2 mm when in fruit; carpels 3; styles 3, 3 mm long, each basally bifurcate, stigmata flabellate sometimes shallowly cupulate; stamens 5, 4 mm long; seeds black, 0.6 mm long, narrowly ovoid-ellipsoid, finely reticulate-foveolate. Roots fine and somewhat succulent.
Drosera camporupestris is included in subgen. Drosera sect. Oospermasensu Schlauer (1996) and is related to D. chrysolepis (Figure 3:2A-F). Drosera camporupestris is distinguished by its short stocky stems; stipules 7-13 mm long, apically fimbriate; only 1-2 (rarely 3) functional leaves; and petioles 25-105 X 0.3-1.5 mm long. Drosera chrysolepis differs in its stems 5-46.5 X 0.1-0.5 cm, thickening towards the apex; stipules only up to 10 mm long, the upper 1/2 to 1/3 divided into 5-9 filaments; 3-13 functional leaves per plant; and petioles only up to 45 mm long.
The only place known where both D. camporupestris and D. chrysolepis occur is the Serra do Cipó. Although never observed growing sympatrically, these two taxa have been found in neighboring habitats only a few meters apart without the presence of intermediates--suggesting there is reproductive incompatibility and supporting their status as separate species.
At the Serra do Cipó both D. camporupestris and D. chrysolepis occur in campo rupestre vegetation, but in somewhat different habitats. Drosera chrysolepis is usually found growing in year-round humid soil among tall grasses in sand mixed with black humus or among short sparse grasses in fine sand with white quartz gravel, but it has also been found in boggy humus-rich soil. Drosera camporupestris is found exclusively in seepage habitats with sandy or humus-rich black soils that are boggy during the summer (wet season) but which often dry up completely during the winter (dry season). Although D. chrysolepis is known to occasionally lose its leaves in the late dry season, D. camporupestris is not deciduous nor does it go dormant.
Drosera camporupestris flowers year round, but is more commonly found in bloom early in the dry season, around June or July. Curiously, the petals of D. camporupestris are sometimes a darker pink-lilac at the base. This is the first record of a New World Drosera species with (at least occasional) distinctly bicolored petals. (However, very weakly bicolored petals of other New World Drosera have been observed, see for example the color plate in Schnell, 2002: p.246.)
Silva & Giulietti (1997) mistook D. camporupestris for toppled D. chrysolepis which had their stems decomposed after being covered by soil. Numerous field observations over a ten year period leave no doubt that the above conclusionis wrong. Buried stem remains were never found on any D. camporupestris and greenhouse germination experiments showed that these two taxa grow true from seed to maturity, maintaining their respective differences such as stem length and leaf number.
Drosera camporupestris is endemic to central Minas Gerais state, southeastern Brazil, where it has been collected on highlands near the towns of Congonhas do Norte (c.1250 m elevation), Diamantina (c.1320 m elevation), Gouvea (c.1260 m elevation), Milho Verde (c.1100 m elevation), as well as on the Serra do Cipó (c.1240-1400 m elevation) (see Figure 5).
An incomplete specimen identified as D. chrysolepis collected on the Cordillera Cóndor in southern Ecuador (near the border with Peru) at 2090 m altitude (Gentry 80306--MO) and examined at SPF is apparently conspecific with D. camporupestris. Yet I hesitate to include it under D. camporupestris since it was collected over 4000 km from any of the known sites in Brazil. Further and more complete specimens from Ecuador would need to be examined first. Another specimen collected at 2500 m altitude on the Cordillera Cóndor (Gentry 80462--MO) is also identified as D. chrysolepis, but was unfortunately not examined by this author. The epithet “camporupestris ” refers to the vegetation typical of the Serra do Cipó and other sandstone highlands of Brazil called “campo rupestre” in which this sundew grows.
Drosera chrysolepis and D. graminifolia (a species closely related to D. camporupestris and D. chrysolepis and also endemic to campos rupestres of Minas Gerais) were initially placed in sect. Drosera, which includes the species of subgen. Drosera with fusiform seeds (Schlauer 1996). A closer analysis of these two species showed that their seeds are clearly more ovoid than fusiform in shape. Therefore, D. chrysolepis and D. graminifolia are here removed from sect. Drosera to sect. Oosperma.
Paratypes-- BRAZIL. Minas Gerais: Congonhas do Norte, Serra da Mangabeira, 23/Sep./1982, Furlan et al. 8464 (SPF); Diamantina, estrada p/ Tapetes Arraiolos, 5/Jun./1998, Rivadavia 751 (SPF); Gouvea, estrada Diamantina—Curvelo, 2/Aug./2002, Rivadavia & Gibson 1398 (SPF); Gouvea, estrada Diamantina-Curvelo, 6/Apr./2003, Rivadavia 1594 (SPF); Jaboticatubas, Serra do Cipó, Fazenda da Serra do Cipó, 26/Feb./1992, Rivadavia 90 (SPF); Jaboticatubas, Serra do Cipó, Fazenda da Serra do Cipó, 5/Jun./1994, Rivadavia 291 (SPF); Jaboticatubas, Serra do Cipó, Fazenda da Serra do Cipó, 6/Jul./1995, Rivadavia 447 (SPF); Jaboticatubas, Serra do Cipó, Fazenda da Serra do Cipó, 24/Feb./1996, Rivadavia & Mullins544 (SPF); Jaboticatubas, Serra do Cipó, alguns km antes do Alto do Palácio, 25/Feb./1997, Rivadavia & Pinheiro 556 (SPF); Jaboticatubas, Serra do Cipó, km 112 ou 113 da estrada p/ C.M.Dentro, 9/Jul./1999, Rivadavia, et al. 1097 (SPF); Jaboticatubas, Serra do Cipó, km 112-113 da estrada p/ C.M.Dentro, 4/Apr./2003, Rivadavia 1542 (SPF); Santa Luzia, Serra do Cipó, km 118, 23/Aug./1933, Mello-Barreto 2433 (BHCB); Santa Luzia, Serra do Cipó, km 122, 14/Apr./1935, Mello-Barreto & Brade 1260 (BHCB) (mixed collection with D. chrysolepis ); Santana do Riacho, Serra do Cipó, s/d, Grandi et al. 1414 (BHCB); Santana do Riacho, Serra do Cipó , 24/Jul./1966, Emygdio et al. 2247 (R); Santana do Riacho , km 109 da estrada L.Santa-C.M.Dentro, 6/Sep ./1980, Forero et al. 7763 (SP); Santana do Riacho , S.do Cipó, 400 m antes da bifurcação p/ M.do Pilar e C.M.Dentro, 22/Sep./1993, Campos & Souza 13394 (SPF); Santana do Riacho , Serra do Cipó, estátua do Juquinha, 26/Sep./2002, Feres et al. 50 (UEC); Serro, estrada Milho verde p/ Diamantina, 6/Apr./2003, Rivadavia 1586 (SPF); Serro, Milho Verde, trilha p/ cachoeira do Arco-Íris, 13/Jan./1998, Chaves s/n (BHCB); Serro, Milho Verde, trilha p/ cachoeira do Arco-Íris, 5/Apr./2003, Rivadavia & Neves 1553 (SPF). Doubtful specimens--ECUADOR. Morona-Santiago: Gualaquiza Cantón, Cordillera del Cóndor, 15 km east of Gualaquiza, 21/Jul./1993, Gentry 80306 (MO); Gualaquiza Cantón, Cordillera del Cóndor , 26/Jul./1993, Gentry 80462 (MO).
Drosera viridis F.Rivadavia, spec. nov. TYPE--BRAZIL. São Paulo: Paranapiacaba, em barranco úmido, junto com D. ascendens A.Saint-Hilaire ao lado da estrada de terra para Paranapiacaba, paralela aos trilhos de trem, 2 km antes da cidade, 2/Feb./1996, Rivadavia & Cardoso 510 (holotype -- SPF). Figure 4:1A-G.
Drosera communis affinis sed robustior, foliis viridibus, petiolis glabris marginibus complanatis exceptis; stylibus rectis, stigmatibus indistinctis; seminibus elongatis rectangularibus.
Perennial herbs with leaves in rosettes flat on the ground or semi-erect in wetter habitats. Stipules 1.5-3 X 0.3-0.5 mm, translucent white to reddish in color, the upper 1/2 to 3/4 divided into 5-9 filaments. Leaves 5-28 mm long, spatulate, entirely green; petioles 1.5-15 X 0.5-1 mm, eglandular-pilose along the margins only, elliptical in cross section but somewhat flattened near the margins; lamina 2-13 X 1-7 mm, obovate-rotundate, glabrous abaxially, adaxially covered with numerous pink or red-tipped retentive glands. Inflorescences 1-3, erect or ascending at the base (rarely bifurcating at the apex), glabrous or nearly so at the base of the scape, glandular-pilose especially towards the apex of the inflorescence, 7.5-30 cm long including scape, with 2-12 flowers each; pedicels 0.5-7 mm long, inserted 2-10 mm apart, glandular-pilose; sepals 5 united at basal 1/4 of length, 2-5 X 0.5-1.5 mm, oblong-ovate, abaxially glandular-pilose; petals 5, 3-6 X 2-3 mm, obovate, light to dark-lilac colored; ovary 1-1.5 mm long at anthesis, globose, trilobed, becoming ellipsoid when in fruit, 3-4 mm long ; carpels 3; styles 3, 1.5-2.5 mm long, erect, each basally bifurcate, crowded over the ovary, stigmata indistinct; stamens 5, 2.5-4 mm long; seeds dark brown, 0.8-1 mm long, rectangular, reticulate. Roots fine and fibrous
Drosera viridis is included in subgen. Drosera sect. Droserasensu Schlauer (1996). It is distinguished from D. communis by its overall larger size; leaves always green, never turning red even when exposed to direct sunlight; eglandular-pubescence present only on the petiole margins (which are flattened in relation to the center of the petiole); indistinct stigmata, and rectangular seeds. Drosera viridis is related to D. communis (Figure 4:2A-G) which differs in its reddish leaves when exposed to sunlight; overall more diminutive size; petioles elliptical in cross section (not flattened on the margins) and eglandular-pilose (more or less) abaxially and adaxially; scapes sharply ascending at base; stigmata bilobed to bifurcate; and seeds narrowly fusiform-cuneate, 1-1.2 mm long.
Drosera viridis is usually found growing in waterlogged habitats on streamsides and in seepages, occasionally submerged in water with only the lamina breaking the surface. It occurs among sparse to dense grasses in white-clayey, reddish-lateritic, or even blackish-brown soil rich in humus. Where D. viridis is sympatric with D. communis, the latter occupies a wider range of habitats, extending further into drier soils. A few weak specimens have been found which may represent hybrids between D. viridis and D. communis. Because both taxa are clearly distinguishable when mature it is suspected that even if they do occasionally cross, the hybrids do not reach maturity or are very rare and infertile.
Drosera viridis has been collected in eastern Paraná and São Paulo as well as central Santa Catarina, growing at 550-1100 m elevation. (Figure 5) Santos (1980) described D. communis from Paraná, Santa Catarina, and Rio Grande do Sul as having petioles glabrous or with only a few hairs on the margins and illustrates rectangular seeds. This strongly suggests that at least one, if not all, of the specimens examined in this study by Santos belong to D. viridis, and not D. communis. Droseraviridis may therefore be widespread in southern Brazil and possibly even in adjacent Argentina, Paraguay, and Uruguay. Like D. communis, D. viridis may be found in flower year-round, apparently without a definite flowering period, although larger numbers of plants are usually found in flower in the wet season from December to March.
The epithet “viridis” refers to the typical green coloration of the plants, even when growing in fully exposed habitats where other Drosera species, including D. communis, commonly acquire a reddish coloration.
Paratypes -- BRAZIL. Paraná: Castro, canyon Guartelá, 22/Apr./1995, Rivadavia, et al. 402 (SPF); Jaguariaíva, rodovia p/ P.Grossa, 12/Jan./1983, Pirani et al. 388 (SP); Jaguariaíva, estrada P.Grossa-Sengés, 23/Apr./1995, Rivadavia 407 (SPF); Jaguariaíva, rio das Mortes, 24/Apr./1995, Rivadavia 409 (SPF); Jaguariaíva, rio Cajurú, 24/Apr./1995, Rivadavia 411 (SPF); Palmeira, rio dos Papagaios, 21/Apr./1995, Rivadavia, et al. 396 (SPF); São Luís do Purunã, estrada Curitiba-P.Grossa, 21/Apr./1995, Rivadavia, et al. 392 (SPF); Sengés, rio Funil, 25/Jan./1993, Rivadavia 171 (SPF). Santa Catarina: Irani, campo de Irani, 15/Dec./1964, Smith & Klein 13959 (R). São Paulo: Cotia, estrada p/ Ibiúna, 13/Feb./1996, Rivadavia, et al. 511 (SPF); Itararé, rio Verde, 24/Apr./1995, Rivadavia 417 (SPF); Itararé, rio Verde, 13/Nov./1994, Souza et al. 7210 (ESA); Mogi das Cruzes, 9/Mar./1999, Rivadavia, et al. 841 (SPF); Paranapiacaba, estrada velha p/ Paranapiacaba, 20/Apr./1991, Rivadavia & Demets 2 (SPF); Paranapiacaba, estrada velha p/ Paranapiacaba, 17/Dec./1992, Rivadavia 156 (SPF); Paranapiacaba, estrada que sai por trás da cidade, 9/Oct./1999, Rivadavia, et al. 1178 (SPF); Paranapiacaba, estrada que sai por trás da cidade, 26/Jan./2003, Rivadavia, et al. 1537 (SPF); Paranapiacaba, estrada de terra paralela aos trilhos de trem, 26/Jan./2003, Rivadavia, et al. 1539 (SPF); Parelheiros, ao lado do trilho de trem, 23/Mar./1995, Rivadavia, et al. 344 (SPF).
Discussion
Due to the fact that many closely related plant species or even genera are capable of hybridizing and often producing fertile offspring, plant taxonomists (unlike animal taxonomists) tend to avoid using reproductive isolation as support for specific rank. This habit is enforced by the sad fact that ecological information is usually not available to plant taxonomists when describing new taxa, many of which are based on herbarium specimens alone. However when such information is available, species rank is often applied to similar plant taxa occurring in the same geographic area and yet maintaining reproductive isolation, and therefore retaining their distinctness despite the opportunity to cross. In counterposition when not enough information is available, subspecies rank is more commonly used when variation is imperfectly known and taxonomists may even assume that subspecies boundaries will be blurred by crossing when there is geographic overlap (Goodall & Marchant, 1996).
Therefore, although reproductive isolation is not commonly employed in plant taxonomy it is believed that this is mostly due to lack of information at the moment of publication. In conjunction with the morphological characters described above, reproductive isolation is felt to support the specific status of D. camporupestris, D. grantsaui, D. tentaculata, and D. viridis, which are seemingly isolated reproductively from each other and from all other Drosera species known to occur sympatrically or in neighbouring habitats. The only natural hybrid known is that between the morphologically very distinct D. grantsaui and D. montana var. tomentosa, but this cross is apparently sterile and not capable of backcrossing with either of the parental taxa with no intergradation having ever been observed.
Further support for the specific status of D. camporupestris, D. grantsaui, D. tentaculata, and especially D. viridis is provided by the shapes of their seeds. Seed shape is considered a very important taxonomic character in the genus Drosera and has been used to separate otherwise very similar species, such as D. esmeraldae (Steyerm.) Maguire & Wurdack and D. capillaris Poir. (Maguire & Wurdack, 1957) or D. peltata Thunb. and D. auriculata Backh. ex Planch. (Diels 1906). More recently, seed shape was used as the decisive character in the description a new species from Brazil, D. graomogolensis T.Silva, distinguishing it from the also Brazilian D. villosa (Silva & Giulietti, 1997).
Acknowledgements
I would like to thank Rolf Grantsau for taxonomic discussions and insightful morphological details revealed by his beautiful botanical drawings; Jan Schlauer for the Latin descriptions and critical review; Allen Lowrie for motivation and for sharing his vast field experience; Fábio Pinheiro, Vítor F.O.Miranda, Marcos R.F.Cardoso, José Maurício Piliackas, the Ono family, Linilson Padovese, Marcelo A.K.Fontana, Mauro Peixoto, Gert Hoogenstrijd, Ed Read, Josef Mullins, Robert Gibson, Matt Hochberg, as well as other friends who helped with field work, cultivation experiments, herbarium information, and endless discussions on Drosera systematics and ecology; and finally the directors, curators, and staff of the institutions visited for the study of Drosera herbarium specimens.
Literature Cited:
Diels, L. 1906, Drosera. In: A.Engler, Das Pflanzenreich 4. 112: 99-100.
Goodall, D. and Marchant, N. 1996, Consistency in taxonomic rank: an example from Drosera. Abstracta Botanica 20(1): 1-15.
Maguire, B. and Wurdack, J.J. 1957. The Botany of the Guayana Highlands – Part II. Mem. New York Bot. Gard. 9(3): 331-336.
Saint-Hilaire, A. 1824 a, Plantes Usuelles des Brasiliens 1 (3): 1-4. Grimbert. Paris.
Saint-Hilaire, A. 1824 b, Droseraceae. Histoire des plantes les plus remarquables du Brésil et du Paraguay 1: 253-271. Paris.
Santos, E. 1980, Droseráceas, in Reitz, R. (ed). Flora Ilustrada Catarinense: 1-23. Herbário Barbosa Rodrigues, Itajaí.
Schlauer, J. 1987, Nomenclatural synopsis of carnivorous phanerogamous plants. Carniv. Pl. Newslett. 15: 59-117.
Schlauer, J. 1996, A Dichotomous key to the genus Drosera L. (Droseraceae). Carniv. Pl. Newslett 25: 67-88.
Schnell, D. 2002, Carnivorous Plants of the United States and Canada. Timber Press, Portland. 468p.
Silva, T.R.S and Giulietti, A.M. 1997, Levantamento das Droseraceae do Brasil. Bol. Bot. Univ. São Paulo 16: 75-105.
Silva, T.R.S and Correa, M.D. 2002, Drosera peruensis (Droseraceae), a new species from Peru. Novon 12(4): 543-545.
Taubert, P. 1893, Plantae Glaziovianae novae vel minus cognitae. Bot. Jahrb. Syst. 17: 502-506.
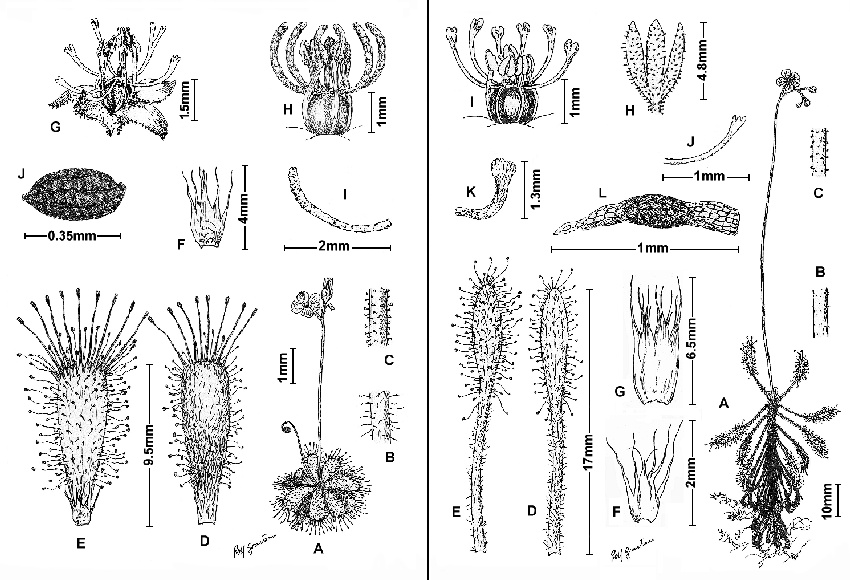
Figure 1 (left): Drosera tentaculata F.Rivadavia showing variation of style morphology from three locations; A: habit; B: lower inflorescence, detail of indumentum; C: upper inflorescence, detail of indumentum; D: leaf, abaxial view; E: leaf, adaxial view with stipule; F: stipules; G: calyx, gynoecium and stamens; H: gynoecium and stamens; I: style; J: seed. (A-F, H, and J based on Rivadavia & Mullins 541 (SPF) from the Serra do Cipó; G based on Rivadavia & Pinheiro 451 (SPF) from Diamantina; I based on Rivadavia 629 (SPF) from the Serra do Cabral.) Figures based on live, pressed and alcohol-preserved specimens.
Figure 2 (right): Drosera grantsaui F.Rivadavia showing variation of style morphology from three locations; A: habit; B: lower inflorescence, detail of indumentum; C: upper inflorescence, detail of indumentum; D: leaf, abaxial view; E: leaf, adaxial view; F and G: stipules; H: calyx; I: gynoecium and stamens; J: style; K: style; L: seed. (A-F, I, and L based on Rivadavia 410 (SPF) from Jaguariaíva; H based on Rivadavia 328 (SPF) from Itacambira; J based on Rivadavia & Cardoso 422 (SPF) from the Chapada dos Guimarães; G and K based on Rivadavia & Ganev 491 (SPF) from the Chapada Diamantina.) Figures based on live, pressed and alcohol-preserved specimens.
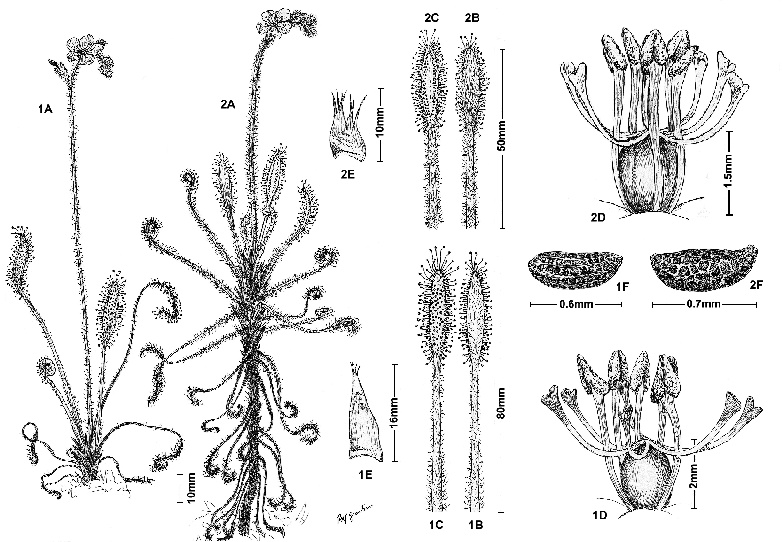
Figure 3: Comparison between (1.) Drosera camporupestris F.Rivadavia (based on Rivadavia 447 (SPF) from the Serra do Cipó) and (2.) D. chrysolepis (based on Rivadavia 212 (SPF) from the Serra do Cipó); A: habit; B: leaf, abaxial view; C: leaf, adaxial view; D: gynoecium and stamens; E: stipules; F: seed. Figures based on live, pressed and alcohol-preserved specimens.
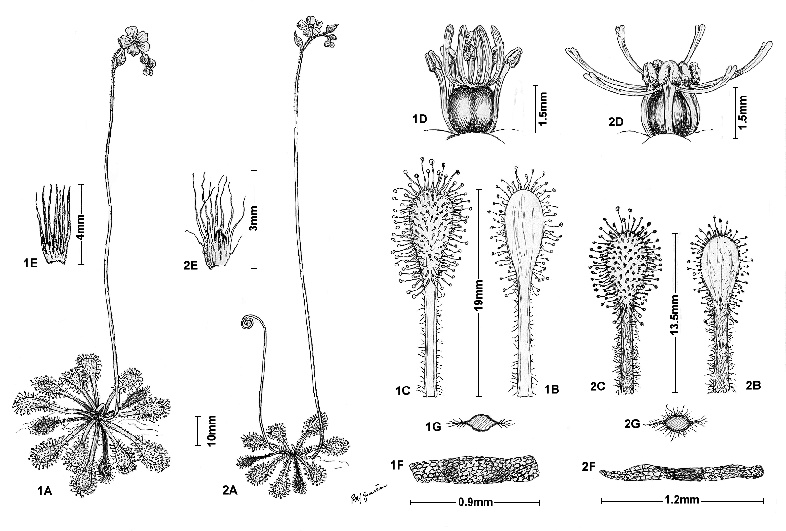
Figure 4: Comparison between (1.) Drosera viridis F.Rivadavia (based on Rivadavia & Cardoso 510 (SPF) from Paranapiacaba) and (2.) D. communis (based on Rivadavia, et al. 343 (SPF) from Parelheiros); A: habit; B: leaf, abaxial view; C: leaf, adaxial view; D: gynoecium and stamens; E: stipules; F: seed; G: petiole in cross section. Figures based on live, pressed and alcohol-preserved specimens.
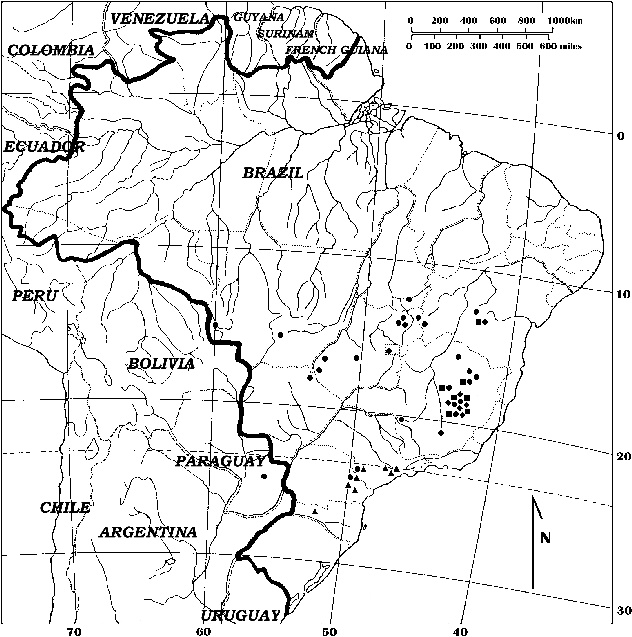
Figure 5: Map showing the known distribution of Drosera tentaculata F.Rivadavia (squares), D. camporupestris F.Rivadavia (diamonds), D. grantsaui F.Rivadavia (circles), and D. viridis F.Rivadavia (triangles). Many of the sites indicated on the map represent collection locations for more than one herbarium specimen listed in this paper.

Figure 6: Drosera camporupestris at Gouvea, Minas Gerais state.

Figure 7: Drosera grantsaui at Itacambira, Minas Gerais state.

Figure 8: Drosera viridis (green plant at center) surrounded by Drosera communis at Paranapiacaba, São Paulo state.

Front Cover: Drosera tentaculata at Gouvea, Minas Gerais state.
|


