|
Carnivorous Plant Newsletter
Volume 37, Number 2, June 2008, pages 36 - 43; Front Cover
Technical Refereed Contribution
The Drosera montana A.Saint-Hilaire
(Droseraceae) complex: A new combination,
Drosera schwackei (Diels) F.Rivadavia, is proposed
Fernando Rivadavia • São Paulo • Brazil
Keywords: New taxa: Drosera montana, Drosera schwackei, Brazil.
Received: 10 July 2007
Abstract
The Drosera montana A.Saint-Hilaire complex is reviewed, discussed and three taxa are excluded from this group: Drosera hirtella A.Saint-Hilaire, D. parvifolia A.Saint-Hilaire, and D. roraimae (Klotzsch ex Diels) Maguire & Laudon. Morphological characteristics, distribution, and habitat preferences, are described for the rare Drosera montana A.Saint-Hilaire var. schwackei Diels. Based on the differences observed between this taxon and other Drosera species native to South America, it is here proposed that it should be elevated to specific status and the new combination D. schwackei (Diels) F.Rivadavia is made.
Introduction
Numerous doubts and changes in taxonomic classifications have plagued nearly 200 years in the history of a group of small rosetted sundews (Droseraceae) from South America, ever since Auguste de Saint-Hilaire (1824a, 1824b) described thirteen Drosera taxa from Brazil, including D. montana A.Saint-Hilaire, D. parvifolia A.Saint-Hilaire, D. tomentosa A.Saint-Hilaire var. tomentosa, D. tomentosa var. glabrata A.Saint-Hilaire, D. hirtella A.Saint-Hilaire var. hirtella, and D. hirtella var. lutescens A.Saint-Hilaire.
Eichler (1872) was the first to suggest alterations in this group, reducing D. tomentosa var. glabrata and D . hirtella var. lutescens to synonymy under D. tomentosa and D. hirtella, respectively. A more drastic reduction was carried out by Diels (1906), who placed D. parvifolia as a synonym of D. montana, while D. tomentosa and D. hirtella were considered varieties of D. montana. Diels also described three new varieties: D. montana var. schwackei Diels, D. m. var. roraimae Klotzsch ex Diels (Diels 1906), and later D. m. var. robusta Diels (Diels 1914). A reversal in taxonomic reductions began with Maguire & Laudon (1957), who proposed the new combination D. roraimae (Klotzsch ex Diels) Maguire & Laudon, with D. m. var. robusta as its synonym.
The last 30 years have seen the most intense nomenclatural action involving taxa of the D. montana complex. Returning in part to the views of Saint-Hilaire, Santos (1989) accepted as valid taxa D . hirtella var. hirtella, D. h. var. lutescens, and D. tomentosa, but not D. tomentosa var. glabrata. Oddly enough, no mention was made of D. montana var. schwackei.
Returning to the views of Diels, Silva (1994) accepted only D. montana var. montana, D. m. var. hirtella, D. m. var. roraimae, D. m. var. schwackei, and D. m. var. tomentosa. However, Silva & Giulietti (1997) once again considered D. roraimae a valid taxon. A new species described by Rivadavia (2003), D. tentaculata F.Rivadavia, is considered a member of the D. montana complex because of the morphological, cytological, and phylogenetic affinities with this group (described below).
In the most recent event of taxonomic reversals involving the D. montana complex, no varieties were recognized by Correa & Silva (2005), who inexplicably lumped as synonyms under
D. montana numerous taxa: D. communis A.Saint-Hilaire var. pauciflora Eichler, D. hirtella var. hirtella, D. h. var. lutescens, D. montana var. schwackei, D. parvifolia, D. tomentosa var. tomentosa, D. t. var. glabrata, and D. pumilla E.Santos.
Most of the taxonomic confusion involving the D. montana complex originated from the excessive taxonomic weight placed on inflorescence indumentum, starting with Diels (1906) in his delimitation of D. montana varieties. Other important characteristics, such as leaf shape, were mostly ignored, possibly because herbarium specimens of small rosetted Drosera species are often badly pressed and rather difficult to study. Diels’ errors were sadly propagated over the past century, helped by the fact that ecological information for the numerous taxa cited above has been mostly inexistent or overlooked. Unfortunately, this lumping reached inexcusable levels with Correa & Silva (2005), whose work will sadly be used as a reference by countless botanists in the future due to the importance of the Flora Neotropica project.
Between 1990 and 2006 nearly all the known Drosera species native to South America were studied extensively in their natural habitats by this author, including all type locations of D. montana complex taxa. Based on new ecological knowledge acquired in the field together with new morphological information obtained from the study of numerous herbarium specimens, including holotypes, a revision of the D. montana complex was deemed necessary, starting with D. m. var. schwackei (see Figures 1, 2, Front Cover). Recent cytological (Rivadavia et al. 2003) and phylogenetic (Rivadavia 2005) data are also here used to define which taxa truly belong in the D. montana complex. The type specimen of D. communis A.Saint-Hilaire var. pauciflora Eichler was examined and is here considered synonymous with D. montana var. montana.
Results
Morphologically, D. montana var. montana and D. m. var. tomentosa are much more similar to each other than either is to D. m. var. schwackei . In agreement with Saint-Hilaire (1824a), D. montana and D. tomentosa are here treated as separate species, and not varieties as proposed by Diels (1906). This will be further discussed in a future paper, presenting morphological and ecological data to support their specific rank.
Drosera montana var. schwackei is one of the rarest Drosera taxa in Brazil, occupying a very specific ecological niche. As a result, it has remained largely understudied and undercollected since its discovery. Since 1997 various naturally occurring populations of this taxon have been discovered and studied by this author in Minas Gerais, southeastern Brazil. The comparison between these new collections with the few previously known specimens of D. montana var. schwackei shows that it is one of the least variable and most easily distinguishable of all Drosera taxa in Brazil. In fact it is so distinct that a new combination is proposed: D. schwackei (Diels) F.Rivadavia. The morphological and ecological characteristics that distinguish D. schwackei are discussed below, with the help of illustrations and a distribution map.
Although historically considered part of the D. montana complex, recent evidence shows that D. roraimae and D. hirtella should be excluded from this group. The chromosome number of 2n=20 was reported for D. roraimae and D. hirtella, while D. montana, D. tomentosa, and D. tentaculata have 2n=40 (Rivadavia 2005). Furthermore, DNA sequence data (Rivadavia et al. 2003) places D. roraimae and D. hirtella on a branch with mostly New World diploid Drosera species, sister to a branch with Brazilian tetraploid (and supposed tetraploid) taxa including D. montana, D. tomentosa, and D. schwackei among others. Morphologically D. roraimae and D. hirtella differ from remaining D. montana complex taxa in having truly spatulate leaves, instead of leaves obovate-cuneate (D. tentaculata), linear-lanceolate to oblongo-lanceolate (D. schwackei), linear-obovate to obovate-spatulate (D. montana), and oblongo-obovate (D. tomentosa). Therefore, both D. roraimae and D. hirtella are here excluded from the D. montana complex. In agreement with Duno de Stefano & Culham (1995), D. parvifolia A.Saint-Hilaire is here considered conspecific with D. communis A.Saint-Hilaire. Diels (1906) placed D. parvifolia as a synonym of D. montana, probably because it has sparse simple hairs near the base of the inflorescence—a characteristic common in D. communis from southeastern Minas Gerais.
Taxonomy
Drosera schwackei (Diels) F.Rivadavia, comb. nov. Das Pflanzenreich 4. 112 (26): 89. 1906. Holotype: Brazil, Minas Gerais, Serra do Cipó, Apr.1892, Schwacke 8234 (B).
Perennial herbs with leaves in rosettes flat on the ground or semi-erect, sometimes forming columns of accumulated dead leaves up to 2 cm high, with 0.5-2 mm long eglandular hairs as well as sessile (or short-stalked) resinous glands distributed on inflorescences from base to apex, pedicels, bracts and sepals abaxially, petioles adaxially and leaves abaxially, the eglandular hairs being especially long and dense towards the base of the petioles abaxially whilst the sessile glands are especially dense on sepals and pedicels and also present on lamina adaxially. Stipules rectangular, membranaceous, 1-4 mm long, 0.8-2 mm wide at the base, translucent bronze-gold in color, the apical 1/2-1/3 divided into several long laciniae. Leaves oblong-oblanceolate, 6-22 mm long, yellow-green in color, eglandular hairs white, longer and denser towards the abaxial base; petioles 1-5 mm long, 0.5-1.7 mm wide; lamina 4-17 mm long, 1.5-4 mm wide, covered with numerous orange-red retentive glands adaxially. Inflorescences erect, 1-2(3) per plant, 6-18 cm long (including scape), yellow-green to a deep red in color, each bearing 1-9 flowers (up to 16 in forked scapes), eglandular hairs white near the base, bronze-gold towards the apex of the inflorescence, 0.2-1 mm long; bracts ovate-filiform, 1-2 inserted between pedicels, 1-3.5 mm long; pedicels 2-6 mm long, inserted 5-15 mm apart from each other, red in color; sepals 5 oblong-ovate 3-8 mm long, 0.7-2 mm wide, united at basal 1/4-1/5 of length, red in color; petals 5, obovate to ellipsoid-obovate, 5-11 mm long, 5-9 mm wide at apex, dark pink-lilac in color; ovary ellipsoid, trilobed to hexalobed, 1-1.5 mm long at anthesis, in fruit becoming ellipsoid, 2-3.5 mm long; carpels 3; styles 3, forked at the base, 2-4 mm long (including stigmas), dark-pink in color, stigmata lobed to flabellate; stamens 5, 2.5-4 mm long, anthers 1-2 mm long; seeds narrowly oblongo-ovate, honeycombed, 0.65-0.8 mm long, 0.2-0.3 mm wide (see Figure 3).
The leaves of D. schwackei are yellow-green in color, but the numerous orange-red retentive glands on the leaves give the rosettes a beautiful golden-orange appearance. The only other New World Drosera taxa known to have similar coloration is D. sessilifolia A.Saint-Hilaire which is easily distinguishable by its annual habit, spatulate-cuneate leaves, 5 carpels, 5 styles, preference for hot lowland savanna areas, and leaf rosettes that often acquire a deep red or wine-red color towards the end of the growing season.
When studying herbarium specimens, the densely eglandular-pilose scapes alone may not be sufficient to distinguish D. schwackei from some hairy forms of D. tomentosa (although the hairs are sparser and longer in the latter). However the color of the specimen may be diagnostic, since the yellowish leaves of D. schwackei become light-brown when dried while the usually red rosettes of D. montana and D. tomentosa become a dark brown to black in color. A closer analysis will reveal other differences such as more robust leaf rosettes and inflorescences in D. schwackei, acute leaf apex in D. schwackei versus obtuse in D. montana and D. tomentosa, abaxial side of leaves more densely eglandular pilose in D. schwackei than in D. montana and most D. tomentosa, and seeds almost double in length and more oblong than ovate in D. schwackei .
The semi-erect leaf rosettes of D. schwackei with linear-lanceolate to oblongo-lanceolate leaves are more similar in appearance to those of D. ascendens A.Saint-Hilaire than to those of D. montana and D. tomentosa which are mostly flat (except the newest emerging leaf), with leaves linear-obovate to obovate-spatulate or oblongo-obovate, respectively. Drosera ascendens and its closer relatives D. villosa A.Saint-Hilaire and D. graomogolensis T.Silva are easily distinguished from D. schwackei by leaves 0.8-13 cm in length; seeds cuneate-fusiform (D. ascendens and D. villosa) or ovoid (D. graomogolensis); and inflorescences 6-46 cm in length, glandular-pilose only or also eglandular-pilose, bearing up to 33 flowers each.
The robust inflorescences of D. schwackei vary from yellow-green to a deep-red in color, with red pedicels and calyx lobes. Together with the relatively large lilac-pink flowers, the inflorescences of D. schwackei resemble most those of D. graminifolia A.Saint-Hilaire, which is easily distinguished by its narrow leaves reaching 34 cm in length; stipules triangular, golden-bronze colored, 10-25 mm long, 10-15 mm wide; and inflorescences with shorter eglandular hairs, often forking more than once, bearing 7-72 flowers. The scapes of D. chrysolepis Taub. and D. camporupestris F.Rivadavia are also eglandular-pilose (although less dense than in D. schwackei), but these species are easily distinguished by their longer spatulate-lanceolate leaves.
The only other Drosera taxon in Brazil south of the Amazon Basin that may have eglandular-pilose inflorescences is D. cayennensis Sagot ex Diels, here including as its synonyms D. colombiana Fernández-Pérez, D. panamensis Correa & A.S.Taylor, D. pumilla, and D. sanariapoana Steyermark. The indumentum of the scapes is very variable for this species and has been one of the main characters used by taxonomists in attempts to separate D. cayennensis into the different taxa here listed as synonyms. While in some populations only eglandular hairs are present on the scapes, others will have glandular hairs only—or even both. One of these D. cayennensis collections with eglandular-pilose indumentum from the Chapada dos Guimarães (Mato Grosso state, western Brazil) was mistakenly identified as D. montana var. schwackei by Silva (1994) and Silva & Giulietti (1997) and as D. montana by Correa & Silva (2005), but is easily differentiated from either by its spatulate-obovate to spatulate-cuneate leaves in flat rosettes and fragile flower scapes usually no longer than 10cm with only 1-5 flowers.
Drosera schwackei flowers from around March to April, which corresponds to the late wet season/early dry season (or late summer/early autumn). This is a rather unusual flowering period among Brazilian Drosera species, especially for a perennial that does not go dormant. The only other taxon that flowers around this same time of year in Brazil south of the Amazon Basin is D. cayennensis Sagot ex Diels. Similar to D. montana and D. hirtella, D. cayennensis also remains dormant as roots in the dry season, but usually flowers and loses its leaf rosettes earlier in the growing season than the former two species.
One of the most unique characteristic of D. schwackei is the habitat in which it grows. It forms small populations in areas of milky-white quartz gravel mixed with fine white sand. This habitat is sparsely vegetated and prone to intense desiccation in the dry season. Although D. schwackei surprisingly does not go dormant, during drier months its rosettes will lose vigor, the leaves become reduced in size, and the retentive glands often lose their mucilage and curl inwards.
Drosera schwackei is only known from the Cadeia do Espinhaço highlands in central and southern Minas Gerais state, Brazil), where it is known to occur around the towns of Congonhas do Norte, Diamantina, and disjunctly near Ouro Preto. The holotype of D. schwackei was collected at the Serra do Cipó, but the exact location on these highlands is not specified (Diels 1906) (see Figure 4). In the Diamantina area this species occurs between 1200-1300m altitude, at Congonhas do Norte at c.1350 m; whilst the elevations for the other D. schwackei sites are not known.
Drosera schwackei belongs in subgen. Drosera sect. Oosperma sensu Schlauer (1996) due to its seeds being more ellipsoid than fusiform. The closest relative of D. schwackei is believed to be D. tentaculata, which is overall less robust; has orange-reddish obovate-cuneate leaves, each with 8-12 distinct, horizontally flat, retentive glands on the apical margin; inflorescences more sparsely eglandular-pilose as well as glandular-pilose; and seeds 0.3-0.45 mm long, ovoid-ellipsoid, reticulate. Although D. tentaculata regularly grows in similar neighboring quartz gravel habitats, it has curiously never been seen growing sympatrically with D. schwackei, indicating differences not yet fully understood in ecology between both taxa. Other Drosera species observed growing near (but never sympatrically with) D. schwackei are D. graminifolia and D. tomentosa.
Specimens Examined—BRAZIL. Minas Gerais: Congonhas do Norte, Serra da Mangabeira, 23/Apr./1982, Furlan et al. 8463 (SPF); Congonhas do Norte, Serra Talhada, 24/Apr./1982, Furlan et al. 8472 (SPF); Congonhas do Norte, S. da Mangabeira, 26/Feb./1997, Rivadavia & Pinheiro 566 (SPF); Diamantina, estr. p/ T.Arraiolos, 27/Feb./1997, Rivadavia & Pinheiro 577 (SPF); Diamantina, estr. p/ T.Arraiolos, 2/Mar./1997, Rivadavia et al. 593 (SPF); Diamantina, estr. p/ T.Arraiolos, 3/Mar./1997, Rivadavia 596 (SPF); Diamantina, estr. p/ T.Arraiolos, 3/Mar./1997, Rivadavia 597 (SPF); Diamantina, estr. p/ T.Arraiolos, 10/Jul./1999, Rivadavia et al. 1105 (SPF); Diamantina, estr. de T.Arraiolos, 31/Jul./2002, Rivadavia & Gibson 1375 (SPF); Diamantina, estr. p/ Cons.Mata, 29/Jun./2003, Rivadavia 1631 (SPF); Diamantina, estr. de T.Arraiolos, 6/Apr./2003, Rivadavia 1588 (SPF); Diamantina, 2km da cidade, 12/May/2007, Rivadavia 2537 (SPF); Ouro Preto, Itacolomi, 25/Jun./1884, Glaziou 14482 (mixed with D. tomentosa) (R); Serra do Cipó, Apr./1892, Schwacke 8234 (OUPR) (Lectotype).
Acknowledgements: I would like to thank Rolf Grantsau for the botanical drawing of D. schwackei; Allen Lowrie and Jan Schlauer for numerous taxonomic discussions over the years; Fabio Pinheiro, Robert Gibson, Marcelo A.K. Fontana, Charles and Jackie Clarke who helped discover and study natural populations of D. schwackei; Barry Rice and Andreas Fleischmann for helpful comments and suggestions; and especially Thomas Carow whose pictures of D. schwackei near Diamantina in 1987 tantalized me for many years, long before I knew what it was, and until I finally rediscovered this rare taxon in nature in 1997. The easily accessible and numerous D. schwackei populations present in a small area near Diamantina were essential in my study of this taxon and were found thanks to a picture taken by Thomas Carow where Diamantina was visible in the background. Thomas took the risk of killing the only surviving D. schwackei in cultivation, by plucking leaves and a flower scape and sending these to me in alcohol for study. I am indebted to Thomas for this sacrifice, especially knowing the anguish it must have caused him. Without the help of Thomas Carow, the present work would not have been possible. No permits were required to conduct the studies described in this paper.
References
Correa, M. and Silva, T.R.S. 2005. Drosera (Droseraceae) in Flora Neotropica Monograph 96. New York Botanical Garden Press.
Diels, L. 1906. Drosera. In A.Engler, Das Pflanzenreich 4. 112: 88-93.
Diels, L. 1914. Droseraceae in Not. Bot. Gart. Berl. 6: 136.
Duno de Stefano, R. and Culham, A. 1995. Dos especies nuevas del género Drosera (Droseraceae) en Venezuela y otros comentarios taxonómicos. Novon 5: 241-245.
Eichler, A.G. 1872. Droseraceae, in Martius, K.F.von, & Eichler, A.G.(eds.). Flora Brasiliensis 14(2): 385-398.
Maguire, B. & Laudon, J.R. 1957. Drosera roraimae. In Maguire & Wurdack, The Botany of the Guayana Highlands Part II. Mem. NY. Bot. Gard. 9(3): 33-34.
Rivadavia, F. 2003. Four new species of sundews, Drosera (Droseraceae), from Brazil. Carniv. Pl. Newslett. 32: 79-92.
Rivadavia, F. 2005. New Chromosome Numbers for Drosera L.(Droseraceae). Carniv. Pl. Newslett. 34: 85-91.
Rivadavia, F., Kondo, K., Kato, M., and Hasebe, H. 2003. Phylogeny of the sundews, Drosera (Droseraceae), based on chloroplast rbcL and nuclear 18S ribosomal DNA sequences. Am. J. Bot. 90(1): 123-130.
Saint-Hilaire, A. 1824a. Plantes usuelles des Brasiliens 1(3): 1-4. Grimbert. Paris.
Saint-Hilaire, A. 1824b. Droseraceae. Histoire des plantes les plus remarquables du Brésil et du Paraguay 1: 253-271. Paris.
Santos, E. 1989. O gênero Drosera L. no Brasil II—sobre D. montana St.Hil. Bradea 5(21): 249-256.
Schlauer, J. 1996. A dichotomous key to the genus Drosera L. (Droseraceae). Carniv. Pl. Newslett. 25: 67-88.
Silva, T.R.S. 1994. Estudos taxonômicos de Drosera L. (Droseraceae) do Brasil. Dissertação de Mestrado. Instituto de Biociências, Universidade de São Paulo, São Paulo.
Silva, T.R.S. and Giulietti, A.M. 1997. Levantamento das Droseraceae do Brasil. Bol. Bot. Univ. São Paulo 16: 75-105.

Figure 1: Drosera schwackei (Diels) F.Rivadavia rosette near Diamantina, Brazil.

Figure 2: Drosera schwackei (Diels) F.Rivadavia plants.
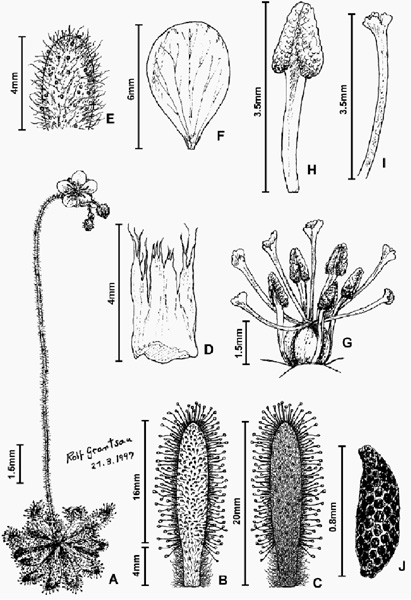
Figure 3: Drosera schwackei (Diels) F.Rivadavia; A: habit; B: leaf, adaxial view; C: leaf, abaxial view; D: stipule; E: sepal, abaxial surface; F: petal, adaxial surface; G: gynoecium and androecium; H: filament and anther; I: a single style/stigma segment; J: seed. Based on live specimens from Congonhas do Norte, Serra da Mangabeira (herbarium specimens from this location deposited at SPF: Rivadavia & Pinheiro 566).
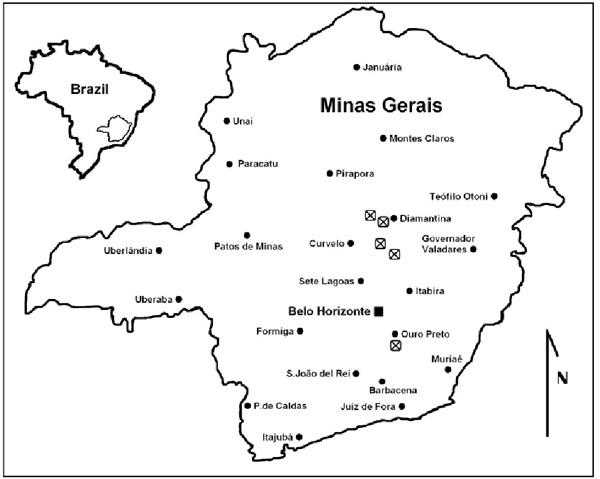
Figure 4: Map showing the known distribution of D. schwackei (Diels) F.Rivadavia. Sites indicated on the map may represent more than one collection listed in this paper. The Serra do Cipó location for Schwacke 8234 (OUPR) is approximated due to the lack of more specific location data.

Front Cover: Newly recognized as a distinct species, Drosera schwackei (Diels) F.Rivadavia, growing on a quartz substrate. Photograph by Fernando Rivadavia.
|


